Research Projects
Behavioural Reproductive Isolation in Drosophila
Multiple types of reproductive barriers have arisen over time to prevent different populations of animals from mating with each other. This keeps different species distinct. One type of barrier is behavioural isolation, which is a prezygotic mechanism that is usually determined by female preference. One way female preference is exhibited is by female fruit flies of one species rejecting males of another species. Drosophila simulans females will reject D. melanogaster males by running away or fighting off any mating advances.
My undergraduate research looked at what genes could be involved with female rejection behaviour towards a male of another species. I used mutant D. melanogaster females that did not express a specific gene and mated them to males of their own species or males of another species to see if they would accept or reject the males. I found that the gene transformer-2 (tra2), a gene involved with the sex determination pathway of Drosophila, could be involved with female rejection behaviour of different species.

The sex determination pathway in Drosophila. In females (left), the Sex lethal (Sxl) gene splices transformer (tra), which works with the protein of transformer-2 (tra2) Tra2 to alternatively splice doublesex (dsx) and fruitless (fru) to give female specific products. In males (right), both male-specific copies of Dsx and Fru are expressed, contributing to male differentiation.
My Master's research investigated the role of the gene fruitless (fru) in behavioural isolation. I aimed to knock-out the gene in both D. melanogaster and D. simulans females using the genetic engineering tool CRISPR. I created a nucleotide deletion in the common region of fruitless, a portion of the gene that is highly conserved amongst different species. This mutation did not have an effect on mating behaviour, suggesting that another coding region of fruitless is involved with female mate preference. You can read my full thesis here.
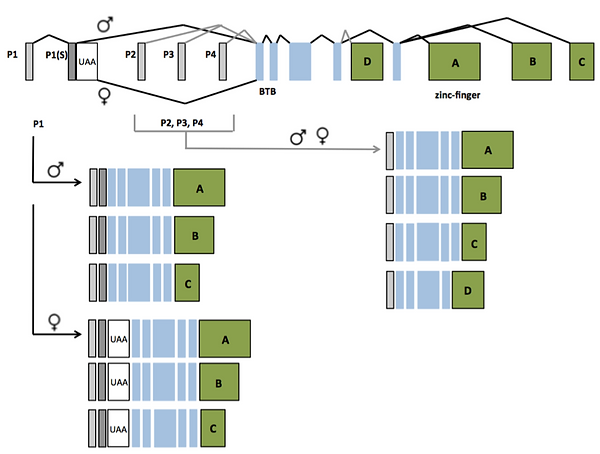
The organization of the fruitless (fru) gene and mRNA products that derive from alternative splicing in males and females. Grey boxes on the 5' end are the four different promoter regions (P1-4), where P1 undergoes alternative splicing in males and females under the control of tra and tra2. Blue boxes are the common region, where the first two exons (boxes) make the BTB protein domain in Fru protein, and the green boxes compose the four different zinc-finger domains. The BTB domain and zinc-finger domains are important for allowing Fru to act as a transcription factor.

Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) is a gene-editing tool adapted from a bacterial immunodefence system. Scientists design sgRNA to be complementary to their sequence of interest so it can bind onto the target sequence. The target sequence must be followed by a PAM sequence (three specific nucleotides) as this helps the Cas9 protein to recognize the sequence it needs to cut. The sgRNA and Cas9 form a complex, allowing for Cas9 to cause a double strand break at the cleavage site (scissors) in the target sequence of interest.
Gametic Reproductive Isolation in Caenorhabditis
When populations of different species come into contact, barriers at the prezygotic stage will be the first to act to reduce gene flow through premating isolation mechanisms such as behavioural and mechanical isolation. If prezygotic barriers cannot prevent copulation between individuals of different species, then gene flow can still be restricted between populations through postzygotic isolation barriers that cause hybrid sterility or inviability. In between these two barriers are postmating isolating barriers, consisting of copulatory behaviour isolation and gametic isolation, that act after sperm transfer but before fertilization. Since postmating isolating barriers act within the reproductive tract, making them difficult to study, they have often been overlooked when identifying prezygotic and postzygotic mechanisms. Therefore, in order to better understand the process of speciation, it is important to understand the dynamics of how postmating isolation barriers act to impede gene flow.
The nematode roundworm Caenorhabditis is great to use to study gametic isolation because it is transparent, meaning that we can see sperm and eggs inside them. They also have ameboid sperm cells that can crawl through the female reproductive tract and break into the female body cavity of other species. This invasive sperm is a form of gametic isolation between species as it can sterilize and/or kill females.
My PhD work was focused on determining the genetic basis of invasive sperm by investigating what genetic components of sperm can make them invasive and how females can become resistant to invasive sperm. I am no longer conducting this research, but it will be explored by other scientists in the Cutter Lab if you're interested!

Head
Tail
Pictured here is a Caenorhabditis elegans hermaphrodite with a specific mutation that affects ovulation. She has been mated with a C. macrosperma male. The red dots are sperm from a C. macrosperma male that has invaded the female gonads and body cavity. Normally sperm should just be found in the uterus and spermathecae (green blobs).
Hybrid Sterility in Caenorhabditis
Hybrid sterility is a form of postzygotic isolation that prevents different species from forming a new population together because their offspring are sterile. Male hybrids of C. remanei and C. latens are sterile due to developmental defects of their gonads. I began to research at what life-stage these developmental defects begin to occur, but alas more research must be done before conclusive results can be shared!
Assessment Strategies for Incorporating Socio-Scientific Issues in the Classsroom
Many Canadian provincial science curriculums incorporate science-in-context frameworks, such as STSE, to help promote global responsibility and agency in their students. These frameworks often encourage students to explore social, technical, cultural, and environmental issues by emphasizing current problems and potential solutions. However, the moral implications and ethical landscapes of these scientific issues are often left unexplored, which means that students are missing out on a key element of learning that will benefit them when faced with a real-life situation where there might not be one perfect solution.
As part of my Master's degree, I conducted qualitative research on this subject by interviewing Canadian educators to better understand what strategies they use to explore moral and ethical issues and be able to assess students in this area. Secondary science teachers often incorporate student experience and interest in various issues as a way to introduce morally complex issues into their classrooms. While teachers often utilize lessons and assessments for or as learning to help students explore social responsibility and moral intelligence, evaluating a student's moral decision is difficult. Often students' growth, measured through pre- and post-reflections, is how teachers tackle the issue of evaluating students on morally complicated issues explored in the curriculum.